Deux élèves étudient le problème du couvercle: pouquoi résonne-t-il lorsque exposé à la lumière d’un flash photographique ?
L’IYPT (à prononcer : aie–why–pi–ti) a pour but de propulser les lycéens dans le monde de la physique expérimentale, de manière ludique et originale, dans l’esprit de la Main à la Pâte ou des Olympiades Française de Physique. Il développe leur sens physique et améliore leurs capacités de communication scientifique. Le tournoi en est à sa 27ème édition. Une trentaine de pays participent aux IYPT, certains de longue date (Russie, Allemagne, Etats-Unis, Australie …), d’autres plus récemment (France, Chine, Nouvelle-Zélande, Iran, Singapour).
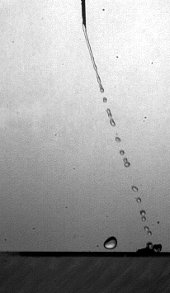
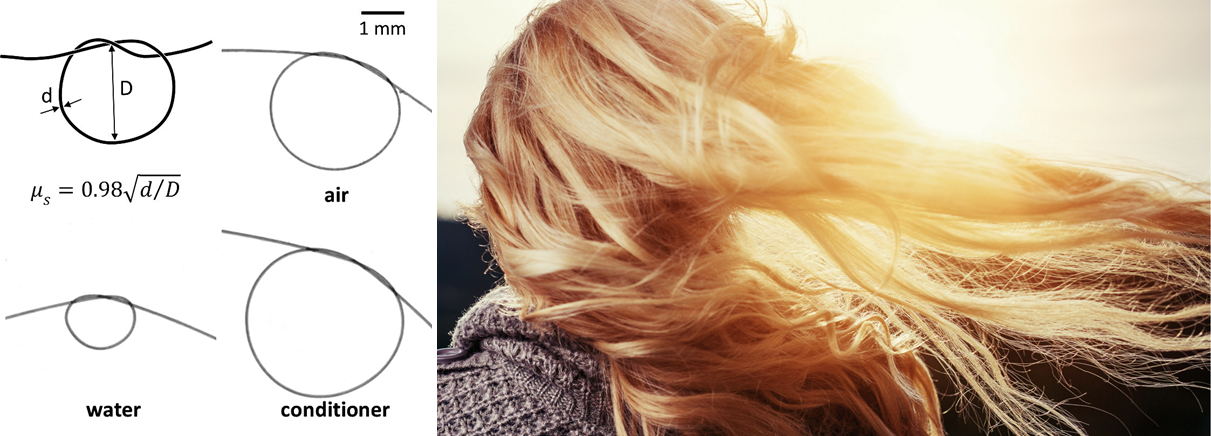
Pourquoi cette goutte rebondit-elle sur une surface hydrophobe ?
Le principe du tournoi est le suivant: chaque été, une liste de 17 problèmes de physique est publiée. Les problèmes sont non-triviaux, à caractère fortement expérimental et ouverts, laissant lieu à des approches et des interprétations différentes. Ils sont le plus souvent inspirés d’observations de la vie courante, par exemple:
- Pourquoi le bord d’une tâche de café séchée est-il plus foncé que sa partie centrale?
- Pourquoi le bord d’une cymbale sonne-t-il lorsque exposé à la lumière d’un flash photographique?
La liste des problèmes 2015 est ici. Chaque équipe sélectionne une dizaine de problèmes parmi ceux proposés et a une année pour les étudier. La résolution des problèmes est en tout point similaire à une activité de recherche scientifique: observation du phénomène, ébahissement, réflexion, expériences plus détaillées, mise en théorie, etc.

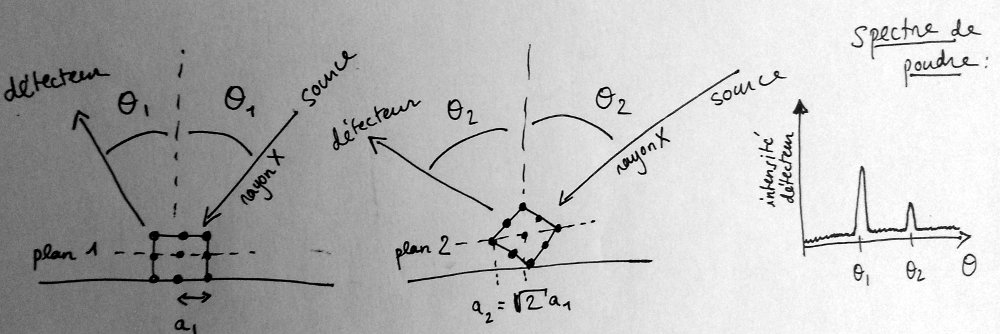
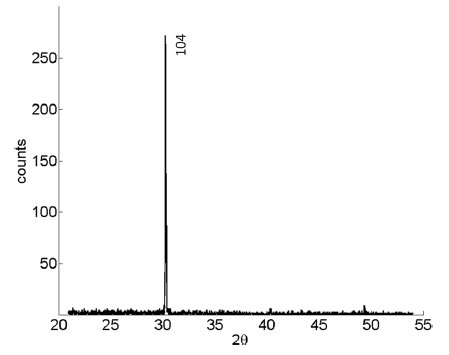
Quelle est l’origine de ses anneaux visibles à l’intérieur d’un tube de cuivre bien poli ?
Le tournoi à proprement parler a lieu chaque année en été dans un pays hôte différent (dernier en date : Taiwan 2013, UK 2014), et dure une semaine. Le tournoi se découpe en débats, au cours desquels les équipes doivent tour à tour exposer leurs solutions, critiquer, arbitrer, tout cela en anglais. Des points sont décernés par un jury composé de scientifiques. L’aspect culturel joue un rôle important lors de la rencontre: des visites sont organisées sur les lieux du tournoi; l’atmosphère détendue se prête idéalement aux échanges entre lycéens de tant de cultures différentes.
L’IYPT en France
Vous trouverez des informations détaillées sur le site : www.iypt-france.org. Contactez-moi directement si ce projet vous interpelle.

Ombre d’un film de savon en lumière laser
J’ai initié avec Gilman Toombes et Patrice Bottineau la première participation française au tournoi de Tianjin en Chine en 2009. Depuis, la France a participé 4 fois, et décroché une médaille de bronze lors du tournoi 2012 de Bad Saulgau en Allemagne.
La préparation scientifique est actuellement gérée par des chercheurs et ingénieurs d’Altran Research et plusieurs ingénieurs de cette entreprise. Elle a lieu tous les mercredi après-midi au lycée Louis-le-Grand, qui nous accueille depuis le début de cette aventure. Les lycéens impliqués (une trentaine chaque année) viennent du tout Paris et de sa banlieue. La préparation est soutenue par plusieurs organismes et entreprises.
L’IYPT a un grand frère, l’IPT, qui suit le même principe mais vise un public d’étudiants d’université, prépa et écoles d’ingénieurs.

L’équipe 2009 en Chine
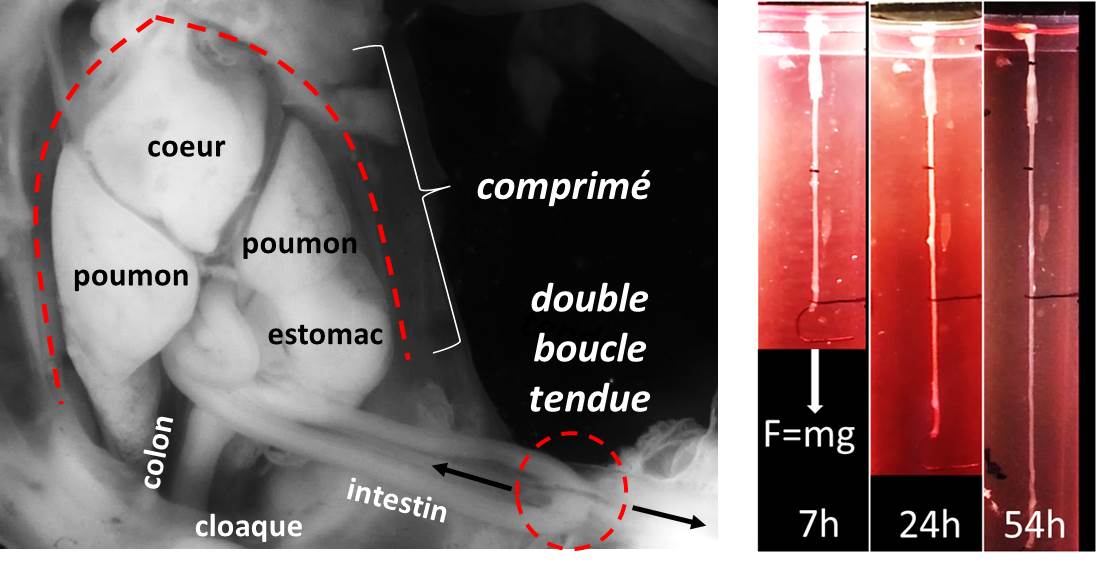
 Un time-lapse sur 6 heures de segments d’intestins oxygénés en permanence. L’agitation est causée par les contractions cycliques des muscles lisses. On voit les intestins s’allonger progressivement: ils poussent, c’est une première en laboratoire.
Un time-lapse sur 6 heures de segments d’intestins oxygénés en permanence. L’agitation est causée par les contractions cycliques des muscles lisses. On voit les intestins s’allonger progressivement: ils poussent, c’est une première en laboratoire.