Neural crest cells are a population of stem cells that colonize organs during the early stages of embryonic development to form multiple derivatives: facial bones, skin pigments, the myelin sheath of the peripheral nervous system, and the entire nervous system of the intestine. These cells recapitulate the behavior of aggressive tumors through their high proliferation and their ability to deform and invade tissues. Defects in neural crest cell migration give rise to congenital pathologies, known as neurocristopathies, the most common of which is Hirschsprung’s disease: incomplete colonization of the intestine results in a lack of nervous system in the colon. This aganglionosis affects 1 in 5,000 births and must be treated surgically at birth. Its causes remain unknown in more than half of cases.
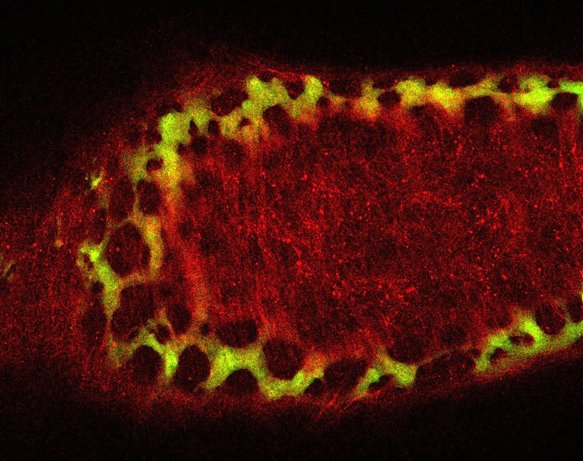
Neural crest cells are in the small intestine and migrate down the cecum and colon, as in viewed in this confocal microscopy time-lapse of an ex-vivo, cultured mouse embryonic gut.
Researchers from the Matter & Complex Systems laboratory (MSC, CNRS, Université Paris Cité), the Imagine Institute (INSERM), and the animal facility of the Institut Jacques Monod have identified a key mechanism that allows neural crest cells to migrate within the intestine. Mutations in a peptide secreted by embryonic intestinal tissue, endothelin 3, are known to cause Hirschsprung’s disease. In adults, endothelin electrically activates the smooth muscles of blood vessels, causing them to constrict, thereby regulating blood pressure. In biology, a protein is said to “moonlight” when, like endothelin, it performs two apparently unrelated functions. The biophysicists at the MSC laboratory did not rely on these appearances: in a study published in the journal Nature Communication, they reveal that endothelin is also the source of electrical oscillations in neural crest cells, and that these oscillations produce the traction force necessary for the cells to squeeze through the developing tissue.
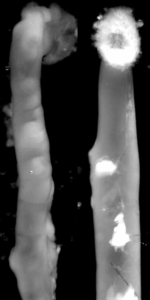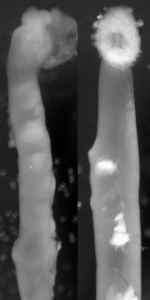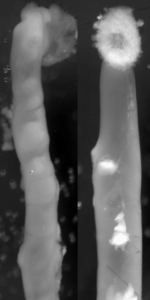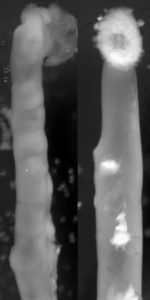
Endothelin 3, a peptide related to Hirschsprung disease, evokes a vivid neural crest calcium response.
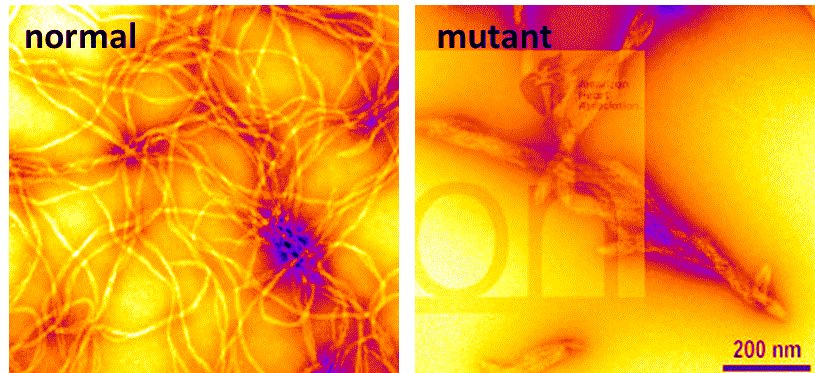
To achieve this result, the researchers used transgenic mouse embryos that fluoresce during calcium oscillations in neural crest cells. They demonstrated that endothelin 3, which is naturally present in embryonic intestinal tissue, drives the oscillations by causing the opening of T-type calcium channels in the membrane. Mutations in these channels have been found in genetic studies of rare variants of Hirschsprung’s disease, although it wasn’t clear at the time why they caused the disease. The researchers then proved, by modulating the degree of opening of the T channels, that the ability of neural crest cells to invade the intestine was entirely linked to their electrical activity. Finally, they developed an ultra-sensitive method to measure the tiny traction forces exerted by neural crest cells on tissue. These cells are not enriched in contractile fibers like the muscle sheath of blood vessels; the researchers therefore placed them in a very soft collagen matrix, studded with nanometric beads to measure the deformation of the medium. The tensile forces exerted by the cells were found to be closely correlated with their calcium activity
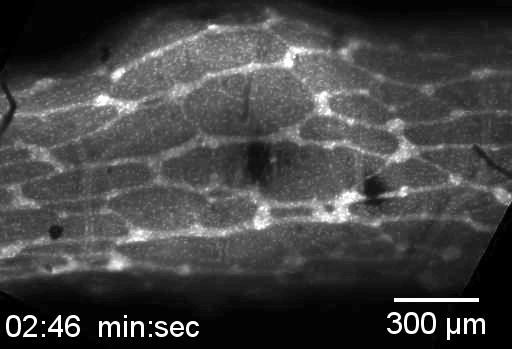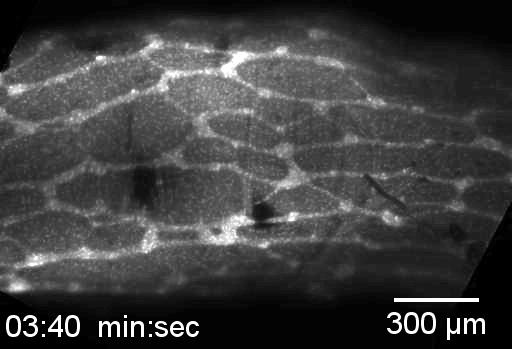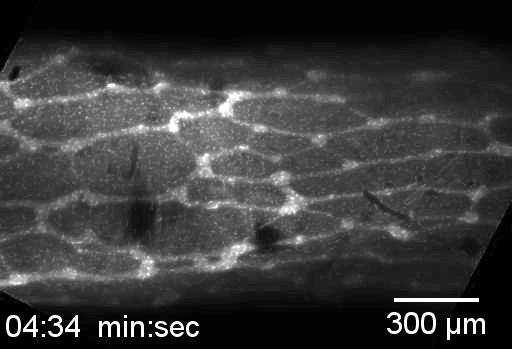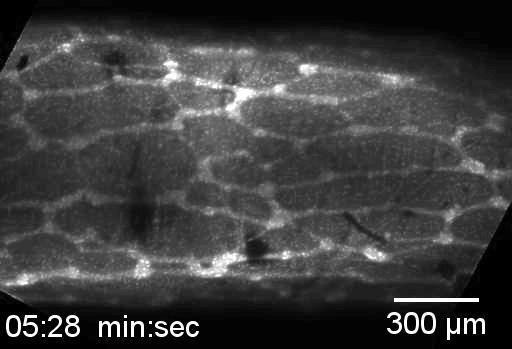
Neural crest cells (in green) are migrating from left to right inside a collagen gel seeded with beads (red). Endothelin 3 puts the gel under tension, because the cells are pulling more. On the contrary, the endothelin 3 receptor inhibitor (BQ788) relaxes this tension.
This study demonstrates that neural crest cells ultimately behave like miniature muscles that contract to colonize the embryo’s organs. This research opens up many possibilities for the diagnosis of neurocristopathies, since a mutation in the factors involved in the electrical or contractile activity of neural crest cells is likely to result in a migration defect, and therefore a pathology. It has also been established that melanomas and neuroglioblastomas result from the reactivation of a neural crest-type program in adults. The migration of these tumors could be prevented by acting on their electrical activity, in particular via T-type calcium channels.
Reference :
Chevalier, N.R., Gayda, F., Bondurand, N. et al. Endothelin-3 and T-type Ca2+ channels drive enteric neural crest cell calcium activity, contractility and migration. Nat Commun 17, 1370 (2026). https://doi.org/10.1038/s41467-025-68121-5